小麦中TAXI-Ⅰ、XIP-Ⅰ和TLXI三类木聚糖酶抑制剂基因检测 
2.安徽省怀远县龙亢农场, 怀远, 233428
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 79 篇 doi: 10.5376/mpb.cn.2011.09.0079
收稿日期: 2011年05月16日 接受日期: 2011年06月15日 发表日期: 2011年06月22日
牛存秀等, 2011, 小麦中TAXI-Ⅰ、XIP-Ⅰ和TLXI三类木聚糖酶抑制剂基因检测, 分子植物育种 Vol.9 No.79 (doi: 10.5376/mpb.cn.2011.09.0079)
木聚糖酶抑制剂基因参与植物的抗病防御反应,在小麦抗病育种中具有一定的应用价值。为了了解木聚糖酶抑制剂基因在不同小麦品种中的分布状况,本文选取来自全国各麦区的260份小麦品种,利用三对特异引物分别对XIP-Ⅰ、TAXI-Ⅰ、TLXI三类木聚糖酶抑制剂基因进行了分子检测,结果表明,在260份材料中,有148份材料含有XIP-Ⅰ基因(占57.0%),116份材料含有TAXI-Ⅰ基因(占44.6%),161份材料含有TLXI基因(占61.9%),60份材料同时含有XIP-Ⅰ和TAXI-Ⅰ两类基因(占23.1%),88份材料同时含有XIP-Ⅰ和TLXI两类基因(占33.8%),75份材料同时含有TAXI-Ⅰ和TLXI两类基因(28.8%),37份材料同时含XIP-Ⅰ、TLXI和TAXI-Ⅰ三类基因(14.2%),20份材料三类基因都不含有(占7.7%)。聚合了三类木聚糖酶抑制剂基因的种质资源可作为抗病育种的亲本。
早在1997年,Debyser等人研究阿拉伯木聚糖的溶解及抑制时发现小麦粉的粗提取物中存在能抑制木聚糖酶活性的蛋白组分(Debyser et al., 1997),并进一步分离纯化,定名为小麦木聚糖酶抑制剂(Triticum aestivum xylanase inhibitor, TAXI) (Debyser et al., 1999)。此后McLauchlan、Fierens等先后从小麦中又分离出两种结构不同的木聚糖酶抑制剂XIP (xylanase inhibitor protein)和TLXI (thaumatin-like xylanase inhibitor) (McLauchlan et al., 1999; Fierens et al., 2007)。迄今为止,在谷物中只发现这三类结构不同的木聚糖酶抑制剂,TAXI和XIP型抑制剂都属多态家族,包含许多结构相近、抑制特异性相似的亚型。后来发现黑麦、大麦、玉米等谷物中也存在TAXI或XIP木聚糖酶抑制剂,推测它们可能广泛存在于单子叶植物中(Goesaert et al., 2004)。许多研究结果表明木聚糖酶抑制剂能抑制病原菌木聚糖酶活性而参与植物的防御反应。
TAXI型木聚糖酶抑制剂属于非糖基蛋白,专一性作用于细菌和真菌的GH11家族木聚糖酶,对GH10家族的木聚糖酶没有抑制活性(Gebruers et al., 2001),目前发现有四种亚型,即TAXI-I、TAXI-II、TAXI-III和TAXI-IV。TAXI-Ⅰ与胡萝卜细胞外周糖蛋白、拟南芥及水稻中的多拷贝序列有较高的相似性,但具不同功能。TAXI-Ⅰ基因的表达受茉莉酸甲酯的强烈诱导(Weng et al., 2010)。TAXI-Ⅲ和TAXI-Ⅳ基因的表达都依赖于病原体和被侵染组织的特异性,在受损叶片中,TAXI-Ⅰ、TAXI-Ⅲ与TAXI-Ⅳ都强烈表达(Igawa et al., 2004)。黑麦和大麦中也存在木聚糖酶抑制剂SCXI和HVXI基因,与TAXI-Ⅰ序列相似(Raedschelders, 2005)。后来许多学者在不同的小麦品种中克隆出TAXI型基因,序列均有差异。
XIP型木聚糖酶抑制剂是一种糖基化单体蛋白,只抑制来自真菌GH10和GH11家族木聚糖酶(Juge et al., 2004)。这种特异性抑制真菌来源木聚糖酶的特征并不是因为真菌酶具有糖基化(Brutus et al., 2004)。研究发现XIP-I具有两个独立的酶结合位点,分别与GH10和GH11家族的木聚糖酶相结合,但有关XIP型木聚糖酶抑制剂的抑制特异性迄今还没有定论。目前发现的有XIP-I、XIP-II、XIP-III、XIP-R1、XIP-R2等亚型。研究XIP-Ⅰ基因表达时发现,XIP-Ⅰ基因在白粉病侵染及受损的叶片中显著表达,被茉莉酸甲酯诱导时表达增强。XIP-Ⅰ基因的表达与病原体的种类和侵染部位有关。XIP-Ⅰ和XIP-Ⅲ基因序列有92.2%的相似性,在功能上XIP-Ⅰ基因起主要作用(Igawa et al., 2005)。Takahashi-Ando等在六倍体小麦的根中又克隆出多条与XIP-Ⅰ型高相似的基因XIP-R,其中XIP-R1为主要成员,在小麦白粉病菌侵染的叶片中表达增强,在赤霉菌感染的小穗中不表达,推测六倍体小麦有一个大的XIP型家族,在特定的组织中仅某些基因表达并参与防御反应(Takahashi-Ando et al., 2007)。
新发现的TLXI型抑制剂,其结构与类奇异果甜蛋白相似,有独特的抑制专一性,以非竞争性抑制方式抑制GH11家族木聚糖酶,同TAXI一样,也不能抑制GH10家族木聚糖酶,属于慢性紧密结合抑制剂(Fierens et al., 2007)。
前人对TAXI-Ⅰ、XIP-Ⅰ和TLXI抑制剂基因在小麦中的分布及其与小麦抗病相关性研究较少。本文选取三对特异引物,采用PCR技术检测260份小麦品种中三类木聚糖酶抑制剂基因,探讨TAXI-Ⅰ、XIP-Ⅰ和TLXI三类木聚糖酶抑制剂基因在不同小麦品种中的存在与分布,以期对小麦抗病基因工程育种做出一定贡献。
1.结果与分析
1.1 XIP-Ⅰ、TAXI-Ⅰ、TLXI三类木聚糖酶抑制蛋白基因检测
1.1.1 XIP-Ⅰ基因检测
用引物对XIP-1F1/ XIP-1R1分别对260份供试育种材料进行XIP-Ⅰ基因克隆,检测结果显示供试品种中57%能扩增出约939 bp的条带,43%的品种没有扩增出任何条带(图1)。琼脂糖凝胶电泳显示,有些品种扩出的条带非常弱,重复几次结果相同,说明该引物较稳定。
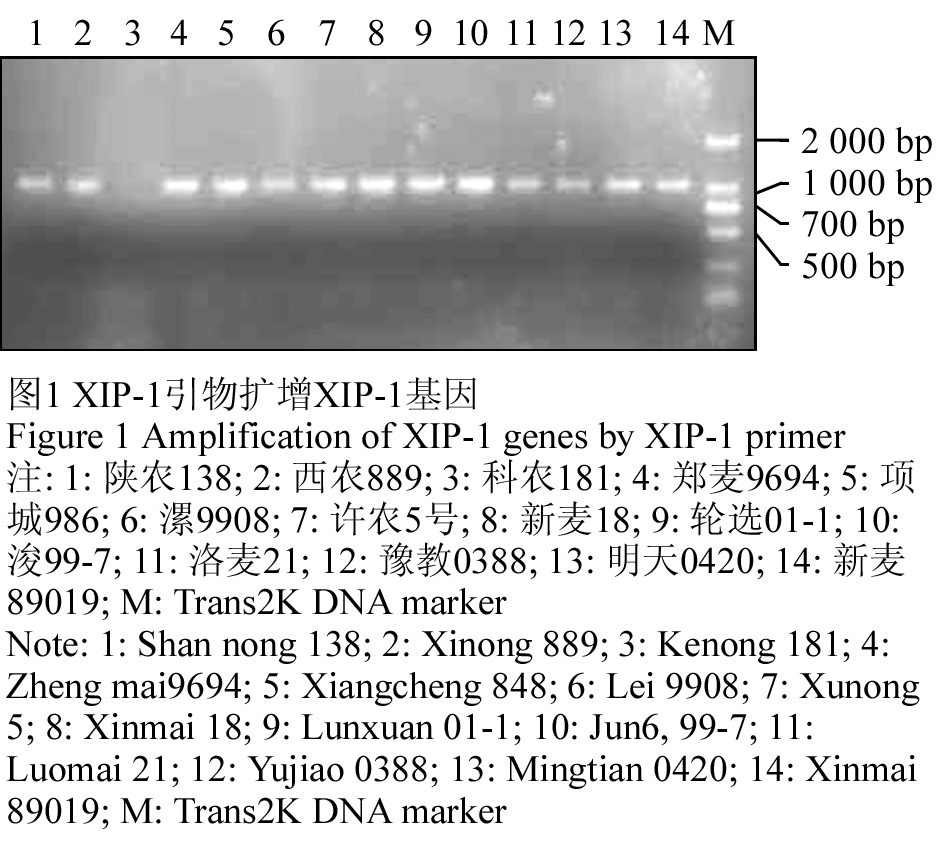
图1 XIP-1引物扩增XIP-1基因
注: 1: 陕农138; 2: 西农889; 3: 科农181; 4: 郑麦9694; 5: 项城986; 6: 漯9908; 7: 许农5号; 8: 新麦18; 9: 轮选01-1; 10: 浚99-7; 11: 洛麦21; 12: 豫教0388; 13: 明天0420; 14: 新麦89019; M: Trans2K DNA marker
Figure 1 Amplification of XIP-1 genes by XIP-1 primer
Note: 1: Shan nong 138; 2: Xinong 889; 3: Kenong 181; 4: Zheng mai9694; 5: Xiangcheng 848; 6: Lei 9908; 7: Xunong 5; 8: Xinmai 18; 9: Lunxuan 01-1; 10: Jun6, 99-7; 11: Luomai 21; 12: Yujiao 0388; 13: Mingtian 0420; 14: Xinmai 89019; M: Trans2K DNA marker
1.1.2 TAXI-Ⅰ基因检测
用引物对TAXI-5/TAXI-3分别对260份供试小麦品种进行TAXI-Ⅰ基因克隆,检测结果显示供试品种中44.6%能扩增出仅有1 256 bp的一条带,其余品种都无扩增条带,约70个品种扩增效果差(图2)。

图2 TAXI-1引物扩增TAXI-1基因
注: 1: 西农4; 2: 西农1; 3: SL02-12; 4: SL02-04; 5: 丰优1718; 6: 丰优161; 7: 旱优63; 8: 旱优62; 9: 旱优61; 10: 登海5348; 11: 登海5049; 12: 烟0469; 13: 烟0428; 14: 泰山6259; M: Trans2K DNA marker
Figure 2 Amplification of TAXI-1 genes by TAXI-1 primer
Note: 1: Xinong 4; 2 Xinong 1; 3: SL02-12; 4: SL02-04; 5: Fenyou1718; 6: Fenyou161; 7: Hanyou 63; 8: Hanyou 62; 9: Hanyou 61; 10: Denghai 5348; 11: Denghai 5049; 12: Yan 0469; 13: Yan 0428; 14: Taishan 6259; M: Trans2K DNA marker
1.1.3 TLXI基因检测
用引物对Lintf/Xi2分别对260份供试小麦品种进行TLXI基因克隆,检测结果显示供试品种中61.9%能扩增出561 bp一条带,其余品种未扩出条带(图3)。

图3 tlxi引物扩增TLXI基因(129-140)
注: 1: 春0501; 2: 新麦9817; 3: 太空6号; 4: 陕715; 5: 内乡203; 6: 西农4211; 7: 春534; 8: 4110; 9: 新原958; 10: 04中36; 11: 丰华8829; 12: 淮核0308; M: Trans2K DNA marker
Figure 3 Amplification of TLXI genes by tlxi primer
Note: 1: Spring 0501; 2: Xinmai9817; 3: Taikong 6; 4: shan 715; 5: NeiXiang 203; 6: Xinong 4211; 7: Spring 534; 8: 4110; 9: Xinyuan 958; 10: 04 zhong 36; 11: Fenghua 8829; 12: Huaihe 0308; M: Trans2K DNA marker
1.2 XIP-Ⅰ、TAXI-Ⅰ、TLXI三类木聚糖酶抑制剂基因在供试材料中的分布
根据上述检测结果,统计XIP-Ⅰ、TAXI-Ⅰ、TLXI三类木聚糖酶抑制剂基因在260份供试小麦品种中的分布。在260份材料中,有35份材料含有XIP-Ⅰ基因,占总材料的13.5%,10份材料含TAXI-Ⅰ单基因,占3.8%,29份材料含有TLXI单基因,占11.2%,60份材料含XIP-Ⅰ和TAXI-Ⅰ,88份含XIP-Ⅰ和TLXI,75份含TAXI-Ⅰ和TLXI,37份含XIP-Ⅰ、TLXI和TAXI-Ⅰ(图4)。
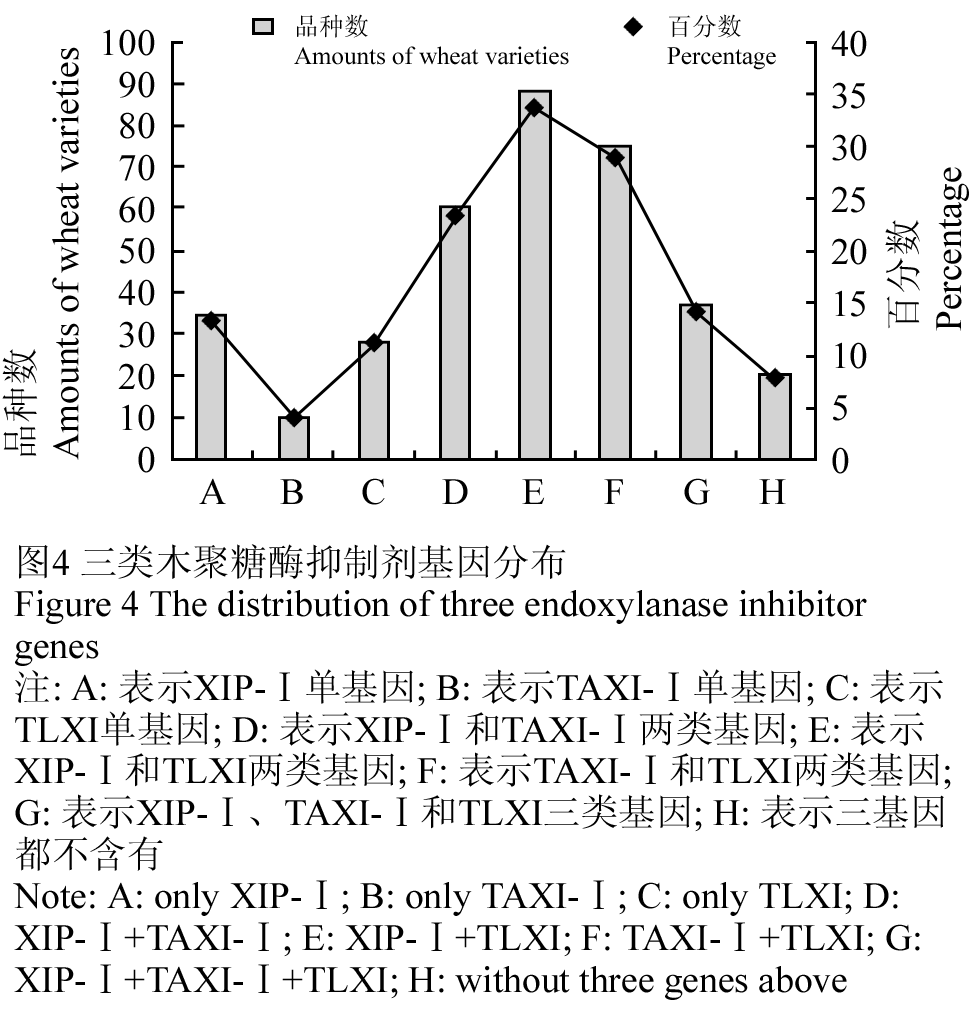
图4 三类木聚糖酶抑制剂基因分布
注: A: 表示XIP-Ⅰ单基因; B: 表示TAXI-Ⅰ单基因; C: 表示TLXI单基因; D: 表示XIP-Ⅰ和TAXI-Ⅰ两类基因; E: 表示XIP-Ⅰ和TLXI两类基因; F: 表示TAXI-Ⅰ和TLXI两类基因; G: 表示XIP-Ⅰ、TAXI-Ⅰ和TLXI三类基因; H: 表示三基因都不含有
Figure 4 The distribution of three endoxylanase inhibitor genes
Note: A: only XIP-Ⅰ; B: only TAXI-Ⅰ; C: only TLXI; D: XIP-Ⅰ+TAXI-Ⅰ; E: XIP-Ⅰ+TLXI; F: TAXI-Ⅰ+TLXI; G: XIP-Ⅰ+TAXI-Ⅰ+TLXI; H: without three genes above
2讨论
本实验对260份不同小麦品种材料的木聚糖酶抑制剂基因检测发现,XIP-Ⅰ、TAXI-Ⅰ、TLXI三类木聚糖酶抑制剂基因在小麦中的分布不同步,且存在差异。有些品种只含有一类基因,有些品种含有两类,有些品种同时含有三类基因,而少数品种不含这些基因。推测在不同小麦品种中木聚糖酶抑制剂基因的类型和序列有差异。
在全基因组中XIP型基因属于大基因家族。Elliott等(2002)最先从小麦品种Soissons中克隆XIP-Ⅰ的cDNA,Igawa等(2005)又从小麦品种Norin 61中克隆出XIP-Ⅲ基因,XIP-Ⅰ和XIP-Ⅲ序列相似度92.2%,但在功能上XIP-Ⅰ基因为XIP型家族的主要基因。Takahashi-Ando等(2007)在六倍体小麦(hexaploid wheat)的根中克隆得到XIP-R1和XIP-R2的cDNA序列,发现在病原防御中XIP-R1基因起主要作用。比对XIP-Ⅰ、XIP-R1 和 XIP-R2基因序列,其相似性较低。推测在不同小麦品种及不同组织中有不同的XIP型基因表达。TAXI型基因家族与XIP型不同,各基因有不同的功能。本实验所检测的三类基因中,TAXI-Ⅰ基因所占比例最少,且扩出的条带较弱,其原因可能是TAXI-Ⅰ型基因序列变异性高。分析NCBI数据库中TAXI-Ⅰ型基因的核苷酸序列发现,同一小麦品种可扩出不同的TAXI-Ⅰ基因序列,例如来自中国春的6条TAXI-Ⅰ序列相似性仅70%左右。
研究表明,XIP型抑制剂与糖苷水解酶18家族的几丁质酶Ⅲ有很高同源性,而TLXI型结构上与类奇异果甜蛋白同源,因此XIP型和TLXI型可分别归为病程相关蛋白(pathogenesis-related proteins) PR5和PR8类型。TAXI-Ⅰ型抑制剂基因的表达受病原体的诱导。已证实三类木聚糖酶抑制剂参与植物的抗病防御反应。本实验只初步分析了不同小麦品种中XIPⅠ、TAXI-Ⅰ、TLXI三类木聚糖酶抑制剂基因的分布,各家族亚型之间的相关性及其与抗病性的关系还需要进一步深入研究。
3材料与方法
3.1试验材料
本实验选用了国内以及国外引种的260份小麦品种进行分子克隆检测,其中包括引种材料35份、地方品种1份、育成或推广品种224份。国内品种/品系分别来自陕西、四川、重庆、河南、山东、河北、安徽、江苏、云南、湖北等多个不同麦区,实验材料具有一定的代表性和广泛性。
3.2试验方法
3.2.1引物的合成
参照Igawa、Fierens等人发表的引物,XIP-1/F1/XIP-1/R1引物对特异扩增XIP-Ⅰ基因序列;TAXI-5/TAXI-3引物对特异扩增TAXI-Ⅰ序列;Lintf/Xi2引物对特异扩增TLXI序列。PCR特异引物均由上海生工生物工程技术服务有限公司合成(见表1)。
![]()
表1 木聚糖酶抑制蛋白基因扩增引物
Table 1 Xylanase inhibiting protein genes amplification primers
3.2.2基因组总DNA的提取
取2粒小麦籽粒磨碎,参照孙道杰博士论文“小麦籽粒PPO活性分子标记及面粉黄色素相关基因研究”中描述的方法(孙道杰, 2005),提取小麦基因组总DNA,用1.2%琼脂糖凝胶电泳检测DNA质量。
3.2.3 PCR检测
PCR扩增于BIO-RAD My Cycler 1.0上进行。反应体系为:总体积10 μL,其中含有0.25 mmol/L dNTP,1×PCR Buffer,2.5 mmol/L Mg2+,上下游引物各0.25 μmol/L,0.5 U DNA Taq聚合酶,40 ng模板DNA。
PCR反应条件为:94℃预变性5 min;94℃变性50 sec,50℃~60℃退火55 sec;72℃延伸l min,共30个循环;最后72℃延伸5 min,4℃保存。PCR扩增产物用2.0%琼脂糖凝胶,以120 V恒压电泳分离,EB染色,凝胶成像系统照相保存。
作者贡献
牛存秀、高雅、陈璨是本研究的实验设计和实验研究的执行人;司红起、牛存秀完成数据分析,论文初稿的写作;高雅、陈璨参与实验设计,试验结果分析;王永玖为本实验提供实验材料;马传喜、司红起是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由现代农业产业技术体系(nycytx-03)和国家科技支撑计划(2009BADA6B01)共同资助,本文中提到了我们实验中涉及的有关试剂供应商和测序服务商,这并非我们为这些试剂供应商和测序服务商的产品和服务提供推荐或背书。
参考文献
Brutus A., Villard C., Durand A., Tahir T., Furniss C., Puigserver A., Juge N., and Giardina T., 2004, The inhibition specificity of recombinant Penicillium funiculosum xylanase B towards wheat proteinaceous inhibitors, Biochim Biophys Acta, 1701(1-2): 121-128
Debyser W., Derdelinckx G., and Delcour J.A., 1997, Arabinoxylan solubilisation and inhibition of the barley malt xylanolytic system by wheat during mashing with wheat wholemeal adjunct: Evidence for a new class of enzyme inhibitors, J. Am. Soc. Brew. Chem., 55: 153-156
Debyser W., Peumans W.J., Van Damme E.J.M., and Delcour J.A., 1999, Triticum aestivum xylanase inhibitor (TAXI-I), a new class of enzyme inhibitor affecting breadmaking performance, J. Cereal Science, 30(1): 39-43
http://dx.doi.org/10.1006/jcrs.1999.0272
Elliott G.O., Hughes R.K., Juge N., Kroon P.A., and Williamson G., 2002, Functional identification of the cDNA coding for a wheat endo-1, 4-β-D-xylanase inhibitor, FEBS Letters, 519(1-3): 66-70
http://dx.doi.org/10.1016/S0014-5793(02)02710-2
Fierens E., Rombouts S., Gebruers K., Goesaert H., Brijs K., Beaugrand J., Volckaert G., Van Campenhout S., Proost P., Courtin C.M., and Delcour J.A., 2007, TLXI, a novel type of xylanase inhibitor from wheat (Triticum aestivum) belonging to the thaumatin family, Biochem J., 403(3): 583-591 http://dx.doi.org/10.1042/BJ20061291 PMid:17269932 PMCid:1876379
Fierens K., Brijs K., Courtin C.M., Gebruers K., Goesaert H., Raedschelders G., Robben J., Van Campenhout S., Volckaert G., and Delcour J.A., 2003, Molecular identification of wheat endoxylanase inhibitor TAXI-I, member of a new class of plant proteins, FEBS Lett, 540(1): 259-263 http://dx.doi.org/10.1016/S0014-5793(03)00276-X
Gebruers K., Debyser W., Goesaert H., Proost P., Van Damm E.J., and Delcour J.A., 2001, Triticum aestivum L.endoxylanase inhibitor (TAXI) consists of two inhibitors, TAXI-I and TAXI-II, with different specificities, Biochem icaf Journaf, 353(2): 239-244
Goesaert H., Elliott G., Kroon P.A., Gebruers K., Courtin C.M., Robben J., Delcour J.A., Juge N., 2004, Occurrence of proteinaceous endoxylanase inhibitors in cereals, Biochim Biophys Acta, 1696(2): 193-202
Igawa T., Ochiai-Fukuda T., Takahashi-Ando N., Ohsato S., Shibata T., Yamaguchi I., and Kimura M., 2004, New TAXI-type xylanase inhibitor genes are inducible by pathogens and wounding in hexaploid wheat, Plant Cell Physiol, 45(10): 1347-1360 http://dx.doi.org/10.1093/pcp/pch195 PMid:15564518
Juge N., Payan F., and Williamson G., 2004, XIP-I, a xylanase inhibitor protein from wheat: A novel protein function, Biochim Biophys Acta, 1696(2): 203-211
McLauchlan W.R., Garcia-Conesa M.T., Williamson G., Roza M., Ravestein P., and Maat J., 1999, A nove1 class of protein from wheat which inhibits xylanases, Biochemicaf Journaf, 338(2): 441-446
http://dx.doi.org/10.1042/0264-6021:3380441 PMid:10024521 PMCid:1220071
Takahashi-Ando N., Inaba M., Ohsato S., Iqawa T., Usami R., and Kimur Ma., 2007, Identification of multiple highly similar XIP-type xylanase inhibitor genes in hexaploid wheat, Biochem Biophys Res Commun, 360(4): 880-884
http://dx.doi.org/10.1016/j.bbrc.2007.06.151 PMid:17631271
Raedschelders G., Fierens K., Sansen S., Rombouts S., Gebruers K., Robben J., Rabijns A., Courtin C.M., Delcour J.A., Van Campenhout S., and Volckaert G., 2005, Molecular identification of wheat endoxylanase inhibitor TAXI-II and the determinants of its inhibition specificity, Biochem Biophys Res. Commun, 335(2): 512-522 http://dx.doi.org/10.1016/j.bbrc.2005.07.103 PMid:16084833
Weng X.Y., Huang Y.Y., Gao H., and Sun J.Y., 2010, Characterization of a xylanase inhibitor TAXI-I from wheat, Biologia Plantarum, 54(1): 154-158 http://dx.doi.org/10.1007/s10535-010-0025-z
Igawa T., Tokai T, Kudo T., Yamaguchi I., and Kimura M., 2005, A wheat xylanase inhibitor gene, Xip-I, but not Taxi-I, is significantly induced by biotic and abiotic signals that trigger plant defense, Biosci. Biotechnol Biochem, 69(5): 1058-1063 http://dx.doi.org/10.1271/bbb.69.1058 PMid:15914935
Sun D.J., 2005, Development of molecular marker for grain PPO activity and detection of flour yellow pigment candidate genes in bread wheat, Dissertation for Ph.D., Northwest A & F University, Supervisor: Wang H., He Z.H., Xia X.,C., pp.27-28 (孙道杰, 2005, 小麦籽粒PPO活性分子标记及面粉黄色素相关基因研究, 博士学位论文, 西北农林科技大学, 导师: 王辉, 夏先春, pp.27-28)